






About Us
Executive Editor:Publishing house "Academy of Natural History"
Editorial Board:
Asgarov S. (Azerbaijan), Alakbarov M. (Azerbaijan), Aliev Z. (Azerbaijan), Babayev N. (Uzbekistan), Chiladze G. (Georgia), Datskovsky I. (Israel), Garbuz I. (Moldova), Gleizer S. (Germany), Ershina A. (Kazakhstan), Kobzev D. (Switzerland), Kohl O. (Germany), Ktshanyan M. (Armenia), Lande D. (Ukraine), Ledvanov M. (Russia), Makats V. (Ukraine), Miletic L. (Serbia), Moskovkin V. (Ukraine), Murzagaliyeva A. (Kazakhstan), Novikov A. (Ukraine), Rahimov R. (Uzbekistan), Romanchuk A. (Ukraine), Shamshiev B. (Kyrgyzstan), Usheva M. (Bulgaria), Vasileva M. (Bulgar).


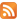
 PDF
PDFNeurotrophicity, neuroprotection and neuroplasticity, along with their connection to morphologic and regenerative processes, play a central role in the structural adaptations of organisms to external influences and compensations that adjust for affected functions.
Results in the last decade have lead to a complete restructuring and reorganisation of the current field and to a changed conception of nervous system function [6].
It is crucial to investigate the biochemical and morphological actions by researching the fundamental and biological process of the nervous system that cause cellular death and are the main targets of therapeutic control.
Neurotrophicity, neuroprotection, and neuroplasticity have no clearly expressed limits. They overlap and mix with each other. Each of them has two aspects: absolute and relative. The absolute side corresponds to mechanisms that cause activation of DNA and result in reparative protein synthesis. The relative side predominantly represents the activation of processes in membranes, the cytosol and cytoplasmic organelles that block cellular death and can induce the appearance of reparative molecules. Absolute mechanisms are predominately controlled by neurotropic factors and by the neurotropic-like molecules. Relative mechanisms are connected with ion channels blockers, agonists, and antagonists of certain receptors by free radical scavengers and metal chelators [5, 6].
Pathologic mechanisms drive different etiological factors or biological instances; therefore, many neurological disorders with different evolutions (acute or chronic) are degenerative. In spite of etiological factors that are not considered homogenous, the processes leading to the disturbance and death are coincident. The aim of neuroprotective treatments consists of blocking these pathogenic processes.
In recent years, no protein has been studied as intensively as P53. Over the last quarter century, it has been the subject of roughly forty thousand scientific research papers, and that number continuing to steadily rise [1]. The protein P53 is not necessary for normal cell differentiation or formation of an organism: the phenotype of nascent mice devoid of the P53 gene differs in no way from normal. Its biological function consists of providing stability of the genome and genetic homogeneity of cells in the whole organism. Therefore, the P53 gene is frequently characterised as the "genome´s guard", "angel-saver", or "gene of cell´s conscience", and mutant genes as "fallen angels" [1, 5, 6].
It has been revealed that disturbances of DNA enable accumulation of P53, which, in turn, blocks progression of the cellular cycle in G1 phase, thereby opposing damage. If reparation of damage is impossible, P53 drives the mechanisms of apoptosis. It is now established that the role of disturbances of the P53 gene in development play a role in not only oncological, but also cardiovascular, neurodegenerative, and metabolic diseases. It is argued that the participation of P53 in the development of autoimmune pathology (Oren M., 2003) would be reflected in the associated processes in the CNS. All of these protective mechanisms could be naturally or pharmacologically active. They are intertwined with each other and together cause complex processes, such as saving and regenerating nerve tissue [1-8].
MATERIALS AND METHODS
Reproduction of the model of ischemic stroke
Experiments were done in male Wistar rats weighting 120-130 g (age 2-3 months), being fed standard rations in a vivarium. Disturbance of cerebral blood circulation was reproduced by a temporary clipping of the left trunk of the anonymous artery; clips were released and the wound closed. Interventions were carried out under general anaesthesia. The animals subject to pharmacotherapy were divided into following groups: Emoxipinum (made in Russia), at 0.1 ml/kg; or placebo (0.9% - 200 Ml solution of NaCl). Control animals were uninfluenced. All animals were randomized and systematized for groups. The efficiency of therapeutic actions was estimated morphologically.
For morphological study of ischemia, we used the pattern that is the widespread model for studying morphological changes of nerve cells after oxygen starvation, the classical levinian preparation (Levine S., 1960). For excluding of influences of the diurnal rhythm, decapitation of animals was carried out under anaesthesia at the same time (9-10 am). Afterwards, the tissue was fixed in 12% solution of formalin and after standard histological manipulation, was filled by paraffin. Coloration of preparations was carried out by hematoxylin eosin, using a standard method. Investigation of histochemical reactions was carried out by means of specified sera of Monoclonal Mouse Bcl-2 and P53 in 50 patterns in polyeosin glasses (MENZEL-GLASER). Photo-optical studies of the investigated material was carried out under a light microscope (Zeiss Axiolab) under passive light in corresponding enlargements; data were saved in a computer database, connected by a special photo and video attachment.
Morphological investigation of temporal lobes after experimental ischemia without treatment
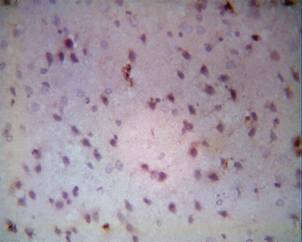
Histological changes in the cortices of the left temporal lobes after experimental ischemia developed over time. Pathological changes in nerve cells after hypoxia were characterised first by polymorphism. Structural changes in nerve cells after hypoxia were detected after 1 hour of chromolysis in different grades in all three investigated groups, to the same degree. Affected neurons start to appear peripherally, centrally or segmentally chromolysed. Comparing patterns in the cortices of the right and left temporal lobes, the primary signs of chromolysis in nerve cells appeared after 60 minutes of restored blood circulation to the left anonymous artery after forcipressure; changes in brains were detected only in some nerve cells and vessels (Fig. 1). The primary signs of brain oedema were characterised by swelling, enlarging in size and bleach-colouring of nerve cells.
|
|
|
|
Figure 1A. The cortex of right temporal lobe after 1 hour of ischemic stroke. Colouring |
Figure 1B. The cortex of the left temporal lobe |
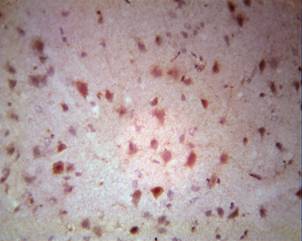
Manifestation of chromatolysis reached a certain intensity after 3-6 hours of occlusion; morphological changes becomes more widespread and abrupt. Acute swelling of the bodies of several neurons and spraying of tigroid and basophilia of the nucleus characterised the further morphological status of the ischemic brain. Chromatolysis, in turn, indicates reactive changes of nerve cells and reflects disturbances in metabolism of functional proteins. In subsequent chromatolysis, the process includes structural proteins of cell, breakdown of which revealed photo-optical incrustation. Products of breakdown of the cytoplasm, intensely-coloured basophilic lumps, developed within 24-36 hours after experiment and finished appearing in cells with the picture of neuronophagia.
At the same time, disturbances of structural proteins and lipids in the cells also include changes to lipoproteins, which testify to deep disturbances in the electrolyte and water balance of the cell. As a result, at earlier stages of the process, chromatolysis often occurs with hydropic changes, which are expressed in vacuolisation of different degrees, enlarge cellular size, and round its cutout. Sometimes it is possible to observe cases of acute or honeycomb swellings of nerve cells.
By investigating the cortex of the temporal lobe with monoclonal tagged Bcl-2 cells over 3-6 hours after ischemia without treatment, light microscopy revealed complex cellular rosetting, in which one neuron was surrounded with multiple Bcl-2 cells (Fig. 2).
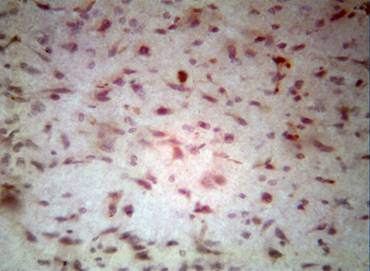
Figure 2. Chaotic allocation Bcl-2 isolated cells around the neurons through 24 hours after ischemia without treatment. Colouring by the hematoxylin-eosin. Enlargement: ob x 40, oc x 10
Chromatolysis and hydropic changes of the cytoplasm at the initial stages, in the first 24-36 hours after adopted hypoxia, is followed by morphological signs of compensating recovery reactions, which are expressed as hypertrophy and hyperchromicity of the ectopic to the nuclear membrane nucleolus (perinuclear hyperchromatosis).
For 24 hours after the experiment, patterns included total hydrolysis and hyperchromatosis of the nucleus in most neurons ("Incrustation" cells, Kariocytolisis, and "Shadow" cells). Some neurons had honeycombed cytoplasm, while others were completely black in colour, with narrow, elongate bodies. Many neurons had central tincturial acidophilia. We observed dystonia of vascular walls, thickening and roughening of the argyrophilic fibres, perevascular swellings and frequently, small perivascular haemorrhages.
Against the homogenic background, a loose basophilic cytoplasm, which appeared darker than at normal, definitely showed a picnomorphic nucleus. Frequently, cells had triangular or rod-shaped forms. The bodies of cells extended, but conserved more or less certain configurations; branches were not coloured (Fig. 3A).
Hypoxic neurons with incrustations were characterised by intensely coloured basophile granules along the periphery of cells, which sometimes formed a darker basophilic borderline in the margin of the cytoplasm.
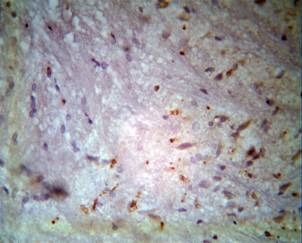
We investigated the cortex by means of monoclonal tagged Bcl-2 cells after 24 hours of ischemia without therapy. Under a light microscope, we observed a chaotic distribution of isolated Bcl-2 cell concentrated around neurons. As a result, we expect further negative dynamics with the appearance of a focal necrosis (Fig. 3).
After 74 hours, the morphological picture changed. On one hand, it was possible to note several signs of restoration: again started colouring powdered to small alpha units - tigroid, and the nuclei of neurons brightened. On the other hand, the pathological proof continued to appear: appearance of vacuolisation, several neurons were darkly coloured, and there was a reduction in sizes of neuronal cells (Fig. 3A).
|
|
|
|
Figure 3A. Depletion by the neuronal cells with the numbers of non-connected Bcl-2 cells of cortex through 3 days after the ischemia without the treatment. Staining by hematoxylin-eosin. |
Figure 3B. The zone of necrotic dissolution of the left temporal lobe of the brain of rats at 7th day after the ischemia without treatment. Independent location Bcl-2 cells. Colouring by the hematoxylin-eosin. Enlarging: ob x 40, oc x 10. |
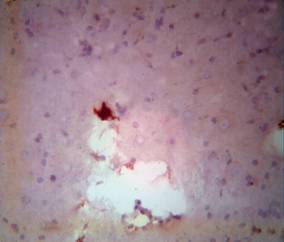
Incrustation, frequently followed by the appearance of different granule cells, was observed with the bleaching of cytoplasm. However, picnotic nuclei could still be detected against the background of the cytoplasm, but gradually converted to a shadow, as an irregularly shaped hyperchrome like decayed to small granules and alpha units of angular formations. Amplification of vacuolisation, with formation of necrotic caves in zones of maximal ischemia and in zones of emptying, characterised morphological processes at the 7th day (Fig. 3B).
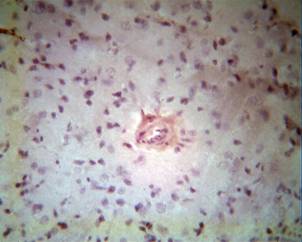
Morphological study of the temporal lobe that was under the influence of emoxipinum following experimental ischemia revealed following peculiarities. We noted, in the populations of nerve cells after 3 hours of emoxipinum exposure, primary cases of chromotolysis that were characterised by a slight brightening of neurons and the appearance of marginal thin bright strips (Fig. 4A). The sizes of cells were relatively stable and were close to normal. Singular neurons were slightly coloured and had round shapes. All current signs were estimated as negligible swellings and persistently demonstrated compensations, appearing as mechanisms of neuroprotection.
|
|
|
|
Figure 4A. Influence of emoxipinum on the cortex of the temporal lobe of the brain of rats, after 3 hours of ischemia. The absence of specific reactions with Bcl-2 cells. Colouring by hematoxylin - eosin. Enlargement: ob x 40, ocu x 10 |
Figure 4B. Singular tags attached to elongated Bcl-2 cells 12 hours after ischemia and treatment by emoxipinum. Colouring with hematoxylin-eosin. Enlargement: ob x 40, oc x 10 |
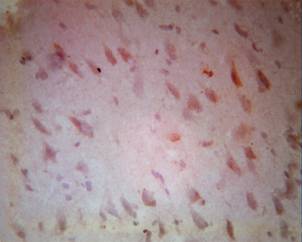
At the interaction of mononuclear tagged Bcl-2 cells with the nerve tissue, we saw a diffusely equal distribution at all surfaces outside of their connections with neurons. As an explanation for this, we found compensatory processes that started within the first hours after the ischemia and activated more-general mechanisms of cellular stability following acute ischemia of the brain.
The following viewpoint on the picture of "uncertainty" could testify to the general reactivity of the nerve tissue. Overall, the receptor mechanism is directed towards the affected cell, and does not influence yet-unstarted necrosis or apoptosis. Such assumptions are not contradictory, but could be proven by the initiation of inflammatory processes and cellular disintegration.
At the 6th hour, we start to see singular cellular exhaustion. In addition, only from this moment does a slight tendency appear for the beginning stages of chromotolysis to be noted. Against this background, there are no tendencies for Вcl-2 cells to group with neurons of the cortex. The areas of ischemic penumbra are relatively narrow. The phenomena of ischemia and perifocal inflammation processes are shown with sufficient expression of glial cells, and are observed especially well in expressed margins of ischemia. An insignificant swelling of neurons, beginning with the manifestation of dissolving of the tigroid and stabilising of processes of chromatolysis, is the distinguishing characteristic after 12 hours of emoxipinum following experimental ischemia (Fig. 4B).
It should be noted that the intensity of chromatolysis is considered to be a neuronal reaction to the metabolism of functionally significant proteins, as the influence of emoxipinum starts to manifest itself directly after the onset of ischemic mechanisms of destruction of brain tissue.
Here, we certify the absence of dissolution of the structural proteins of cells, which manifests under light microscopy as incrustations in the untreated group.
We observed singular tags attached to Bcl-2 cells, against a background of "empty" zones and a tendency for groupings of neuronal cells. The manifestation of such a reaction has been estimated as the primary inclusion of the proliferating mechanisms following processes affecting neuronal cells, which demonstrates not only specific reactions with Bcl-2 cells, but also morphologic manifestations in the structures of brain tissue.
Despite several positive morphologic dynamics at 12th hour following emoxipinum exposure, pathological manifestations of the initial mechanisms of ischemia and hypoxia of the brain also developed further.
In morphological investigations of patterns of the cortex 24 hours after ischemia and treatment with emoxipinum (Fig. 5A), we saw nerve cells areas that were surrounded with enlarged and swollen oxyphil - pyramidal cells with smeared cellular margins, along which there were many polygonal cells. All cells were elongated along the proper axis. Over 3 days after the experiment, changes in the morphological picture of the cortex of the temporal lobe were differentiated by negative dynamics. The pathologic reactions continued to appear: manifestations of empty zones, abruption, and dissolution of neuroglia.
We saw neurons that were elongated, a heavily swollen blurriness of cellular margins, and also observed turned-down and twisted cells. Some cytoplasmic vacuolization hyperchromatosis matched with homogenisations of cytoplasm, and decreed a reduction in sizes of neuronal cells (Fig. 5A). Manifestation of the specific immunohistochemical reactions of Bcl-2 cells were observed in specific surrounds of neurons, where the processes affecting the cells began.
At the 7th day, we observed a relative stabilisation of the processes. Areas that lost nerve cells interchanged with large vacuoles and broad areas of dissolved paths that penetrated the brain tissue (Fig. 5B). Brain cells were less swollen, but hypochromatosis and intracytoplasmic vacuolisation demonstrated a continuing pathological process.
The reaction with tagged Bcl-2 cells begins to manifest its specificity. In the field of view, we saw groupings of tagged Bcl-2 cells directs towards destabilised neurons. There was a tendency for an absence of tagged Bcl-2 cells in some empty areas, around the vacuoles, and along the broad areas of dissolved paths.
|
|
|
|
Figure 5A. Influence of emoxipinum on the cortex of temporal lobe 24 hours after ischemia. |
Figure 5B. The tendency for the specific reaction with Bcl-2 cells rosetting. The absence of Bcl-2 cells in zones emptying at the 7th day after the ischemia |
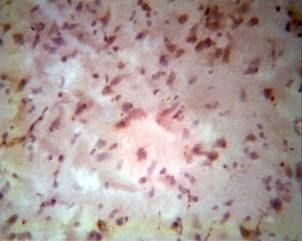
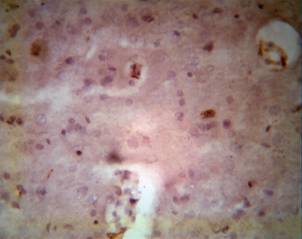
SUMMARY
The morphological and histochemical investigation of neurotrophicity and neuroplasticity after ischemic accidents of the CNS, by means of Bcl-2 and P53, must consider the main marker in early manifestations of the dynamics of the pathological processes. The protective mechanism of the CNS more strongly activates and brings major effects by interaction of natural and pharmacological processes. Of course, such complex and expensive morphological approaches are inconceivable in practical and routine clinical use. We consider that in fundamental studies of the peculiarities of CNS during ischemic accidents, and especially for attempting pharmacological protection and treatment, such investigations are impossible for making a decision.
REFERENCES
- Chumakov PM: Function of gen p53: choice between the life and death / Biochem 2000;65 (1):34-47.
- Iadecola C, Niwa S, Nogawa S, Zhao X, Nagayama M, Araki E et al.: Reduced susceptibility to ischemic brain injury and N-methyl-D-aspartate-mediated neurotoxicity in cyclooxygenase-2-deficient mice / Proc Natl Acad Sci USA 2001;98:1294-99.
- Jun Li, Xibin Liang, Qian Wang, Richard M. Breyer, Louise McCullough, Katrin Andreasson: Misoprostol, an anti-ulcer agent and PGE2 receptor agonist, protects against cerebral ischemia / Neurosci Lett 2008;438(2):210-15.
- Lipton SA: Paradigm shift in neuroprotection by NMDA receptor blockade: memantine and beyond / Nat Rev 2006;5:161-70.
- Myron DG: Neuroprotection for ischemic stroke: past, present and future / Neuropharm 2008;55(3):363-89.
- Sloviter R: Apoptosis: a guide for perplexed / Trends Pharmacol Sci 2002;23:19-24.
- Vecsei L: Kynurenines in the brain: from experiments to clinics / NY: Nova Science Publishers, 2005.
- Zhang ZG, Zhang L, Jiang Q, Zhang R, Davies K, Powers C, et al.: VEGF enhances angiogenesis and promotes blood-brain barrier leakage in the ischemic brain / J Clin Invest 2000;106:829-38.
D. Sagatov, H.Rasulova and Y.Madjidova MORPHOLOGICAL INSIGHT INTO NEW OPPORTUNITIES USING BCL-2 AND P53 IN STUDIES OF NEUROTROPHICITY, NEUROPROTECTION AND NEUROPLASTICITY IN BRAIN ISCHEMIA. International Journal Of Applied And Fundamental Research. – 2010. – № 4 –
URL: www.science-sd.com/386-23436 (03.10.2025).